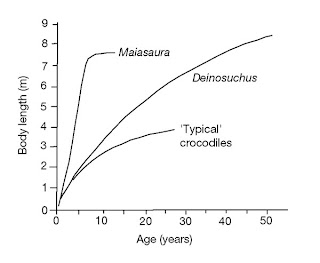
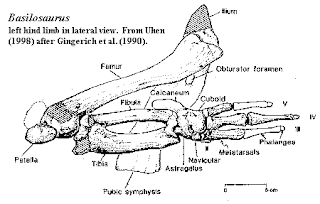
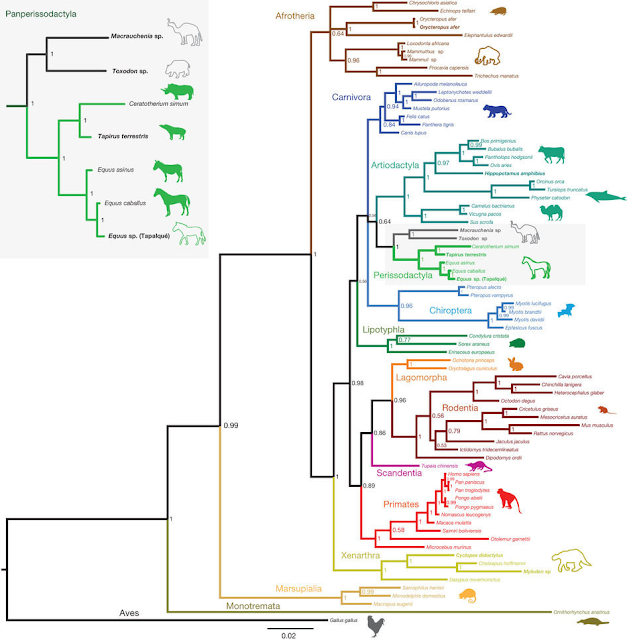
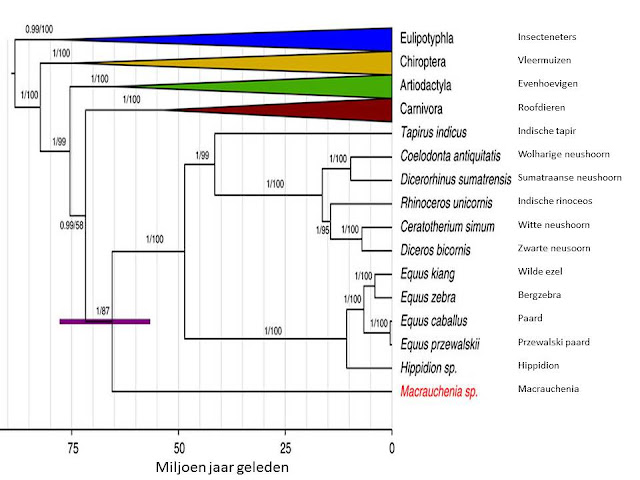
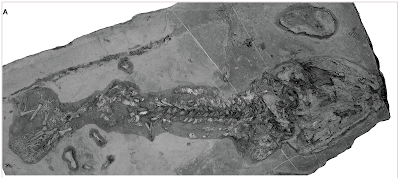
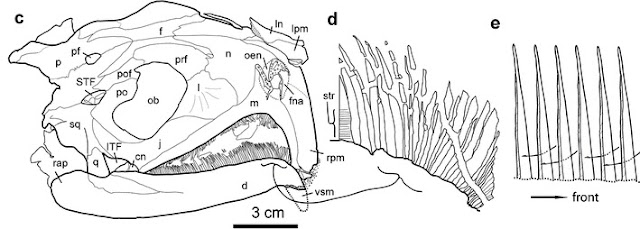
O frabjous day! Callooh! Callay!'
He chortled in his joy.
(Uit: Jabberwocky)
Dit vond ik vrijdag 9 maart heel toepasselijk. Het was het idee van vreugde. Hierom:
![]()
Microraptor in al zijn pracht! Zwart met weerschijn.
Microraptor leefde in het Beneden-Krijt , zo’n 120 miljoen jaar geleden. Er zijn relatief veel en goede fossielen van gevonden. Zoveel goede fossielen dat het verenkleed en de kleur gereconstrueerd konden worden.
Voor de reconstructie werd voornamelijk het fossiele exemplaar BMNHC PH881 (te vinden in het Beijing Museum of Natural History) gebruikt. Bij BMNHC PH881 is bijna het hele skelet aanwezig, in het oorspronkelijke verband, en zijn de veren op voor- en achterpoot en bij de staart duidelijk zichtbaar. Het is een vrij klein exemplaar voor Microraptor. Het beest was ongeveer 54 cm lang van de voorkant van zijn bek tot het einde van zijn staartveren, met een spanwijdte van omstreeks 50 cm (op grond van het meetschaaltje in de figuur). Om een idee te geven: een kauw voor een kleine Microraptor tot een raaf voor een grote Microraptor, met een veel langere staart en een kleinere spanwijdte.
![]()
Microraptor exemplaar BMNHC PH881.
A: foto van het fossiel
B: tekening waarin botten (blauw) en veren (oranje) aangegeven zijn, met de 20 monsterplaatsen voor de studie van de veerstructuur.
Het hele verendek van Microraptor is gereconstrueerd, aan de hand van dit fossiel en een aantal andere fossielen..
![]()
Staartuitende BMNHC PH881
![]()
De staart eindigde in een soort pluim bekroond met twee veren die veel langer zijn dan de andere veren.
![]()
Achterpoten met lange veren BMNHC PH881
De beide achterpoten hebben lange veren aan onderbeen en middenvoetsbeenderen. De tenen steken buiten de veren uit. Voor de voet is dat te zien aan de linkerachterpoot (bovenste) en voor het onderbeen is het te zien aan de rechterachterpoot (bovenste).
![]()
Asymmetrie in de veren van de linkerachterpoot die onder de staart liggen BMNHC PH881
De veren van de voet zijn lang en hebben de vorm van slagpennen. Deze veren zijn duidelijk asymmetrisch, wat erop wijst dat ze van belang zijn voor vliegen.
Bij een vogel – een huidige vogel – zit het verendek van de vleugel, de voorpoot, als volgt in elkaar:
![]()
1. handpennen = primaire slagpennen = grote slagpennen
2. handdekveren
3. duimvleugel
4. armslagpennen = secundaire slagpennen = kleine slagpennen = onderarmpennen
5. grote dekveren
6. middelste dekveren
7. kleine dekveren
8. binnenste armpennen = tertiaire slagpennen = bovenarmpennen
9. schouderveren
Bij Microraptor lijkt de bevedering erg vergelijkbaar met die bij de huidige vogels. Er zijn foto’s van de veren van twee exemplaren, BMNHC PH881 en IVPP V 13352
![]()
De rechtervoorpoot van Microraptor. In de linkerfiguur is de omtrek van botten en slagpennen ingetekend. Er zijn duidelijke handpennen (primaries) en wat minder duidelijke onderarmpennnen (secundaries). BMNHC PH881
Van de rechtervoorpoot van Microraptor zijn bovenarm, spaakbeen, ellepijp en de handbotten goed zichtbaar. Er zijn drie klauwen die buiten de veren uitstaken. De hand heeft duidelijk grote slagpennen, asymmetrisch van vorm. Het aantal slagpennen is zeker groter dan zeven, en het lijkt op 10 zichtbare handpennen. Bij de huidige vogels is het aantal handpennen 9-11.
Bij een eerder gevonden exemplaar zijn de onderarmpennen duidelijker, en zijn ook dekveren zichtbaar. Hoeveel lagen dekveren er waren is niet te zien.
![]()
De rechtervoorpoot van Microraptor. Er zijn duidelijke handpennen (primaries) en duidelijke onderarmpennnen (secundaries). Ook zijn dekveren te zien (coverts). IVPP V 13352.
Kon Microraptor vliegen? Zeker: met vier vleugels. De asymmetrie in de slagpennen van voorpoot en achterpoot laat dat zien. Vliegen als bij een dubbeldekker (als dat zo heet), eerste vliegende versie van het vliegtuig.
De kleur van het verendek van Microraptor kan met goede argumenten worden weergegeven als zwart met weerschijn. Dat gaat op dezelfde manier als de kleur van Anchiornis is beargumenteerd.
![]()
Anchiornis huxleyi was grijs met zwart en witte strepen en een bruinrode kuif.
Al het zwart en grijs bij vogels is het gevolg van een zwarte kleurstof, eumelanine. Bruinig, rossig, gelig of crème is het gevolg van een roodbruine kleurstof, feomelanine.
Melanine zit in een veer opgeslagen in melanosomen. Eumelanine in eumelanosomen; feomelanine in feomelanosomen. Hoe meer melanosomen, hoe donkerder. De vorm van de melanosomen kan gezien worden met behulp van een Scanning Electron Microscope (SEM). Bij de huidige vogels hebben eumelanosomen de vorm van een afgeronde staaf; feomelanosomen zijn meer eivormig.
De melanosomen in de veren van Microraptor bleken erg lang en heel dicht op elkaar gepakt in lagen. Dat deed aan iets denken: aan weerschijn. Weerschijn is het gevolg van de structuur van een stof, met lagen waartussen lichtgolven breken en met elkaar interfereren.
Li, Vinther en anderen keken naar de melanosomen in veren van een aantal vogelsoorten. In totaal zijn er melanosomen van168 soorten bekeken, over bijna alle vogelordes.
![]()
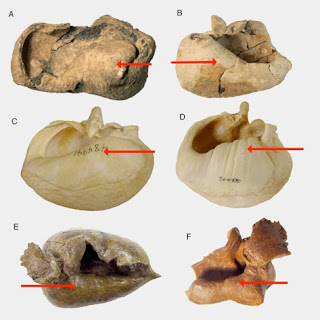
Voorbeelden van de vorm van melanosomen. Melanosomen van (A) Noord-Amerikaanse kuifmees, Baeolophus bicolor, bruin; (B) macaroni penguin, Eudyptes chrysolophus, penguin bruin-zwart; (C) aalscholver Phalacrocorax auritus (grijs); (D) palm kakatoe, Probosciger aterrimus, zwart ; (E) Brazilie eend, Amazonetta brasiliensis, weerschijn; (F) Microraptor, monsterplaats op de schouder.
![]()
De lengte en de vorm van de melanosomen kunnen statistisch worden bewerkt en dan uitgezet in het volgende plaatje.
Statistiek op melanosoom vorm (x-as, Canonical 1) en lengte (y-as, Canonical 2). Stippen horen bij monsters van melanosomen van huidige vogelsoorten. Kleurcode: bruin = bruin, penguin = blauw, grijs = grijs, zwart = zwart, weerschijn = paars.
Nummers geven de Microraptor monsters weer. De Microraptor monsters komen terecht tussen de zwarte en weerschijn punten van de huidige vogels.
Er zijn 20 monsterpunten waar veren van Microraptor herkenbare melanosomen opleverden; de monsterpunten staan in het tweede plaatje boven aangegeven. De Microraptor monsters komen terecht tussen de zwarte en weerschijn punten van de huidige vogels.
Veren met weerschijn hebben langere en smallere melanosomen dan zwarte veren, en melanosomen voor alle andere kleuren zijn boller. Bovendien zijn melanosomen in veren met weerschijn dichter gepakt.
![]()
Tabel die de vorm van de melanosomen laat zien.
Al met al: een zwarte Microraptor met weerschijn.
![]()
Microraptor in al zijn pracht! Zwart met weerschijn.
En als de twee lange middenveren in de staart alleen bij mannetjes voorkomen, is dit een paartje:
![]()
******************
Li, Gao, Meng, Clarke, Shawkey, D’Alba, Pei, Ellison, Norell & Vinther. 2012. Reconstruction of Microraptor and the Evolution of Iridescent Plumage. Science 335:1215-1219. http://dx.doi.org/10.1126/science.1213780
Images: reconstruction by Jason Brougham; fossil photos by Mick Ellison
http://blogs.discovermagazine.com/notrocketscience/2012/03/08/a-shiny-dinosaur-%e2%80%93four-winged-microraptor-gets-colour-and-gloss/
http://en.wikipedia.org/wiki/Microraptor
http://nl.wikipedia.org/wiki/Microraptor
http://evolutiebiologie.blogspot.com/2010/02/sommige-kleuren-van-de-dino.html
http://blogs.discovermagazine.com/notrocketscience/2011/11/21/microraptor-%e2%80%93-the-four-winged-dinosaur-that-ate-birds/
http://blogs.discovermagazine.com/notrocketscience/2008/10/08/microraptor-the-dinosaur-that-flew-like-a-biplane/
http://www.jabberwocky.com/carroll/jabber/jabberwocky.html
http://nl.wikipedia.org/wiki/Iriseren
He chortled in his joy.
(Uit: Jabberwocky)
Dit vond ik vrijdag 9 maart heel toepasselijk. Het was het idee van vreugde. Hierom:

Microraptor in al zijn pracht! Zwart met weerschijn.
Microraptor leefde in het Beneden-Krijt , zo’n 120 miljoen jaar geleden. Er zijn relatief veel en goede fossielen van gevonden. Zoveel goede fossielen dat het verenkleed en de kleur gereconstrueerd konden worden.
Voor de reconstructie werd voornamelijk het fossiele exemplaar BMNHC PH881 (te vinden in het Beijing Museum of Natural History) gebruikt. Bij BMNHC PH881 is bijna het hele skelet aanwezig, in het oorspronkelijke verband, en zijn de veren op voor- en achterpoot en bij de staart duidelijk zichtbaar. Het is een vrij klein exemplaar voor Microraptor. Het beest was ongeveer 54 cm lang van de voorkant van zijn bek tot het einde van zijn staartveren, met een spanwijdte van omstreeks 50 cm (op grond van het meetschaaltje in de figuur). Om een idee te geven: een kauw voor een kleine Microraptor tot een raaf voor een grote Microraptor, met een veel langere staart en een kleinere spanwijdte.

Microraptor exemplaar BMNHC PH881.
A: foto van het fossiel
B: tekening waarin botten (blauw) en veren (oranje) aangegeven zijn, met de 20 monsterplaatsen voor de studie van de veerstructuur.
Het hele verendek van Microraptor is gereconstrueerd, aan de hand van dit fossiel en een aantal andere fossielen..

Staartuitende BMNHC PH881

De staart eindigde in een soort pluim bekroond met twee veren die veel langer zijn dan de andere veren.

Achterpoten met lange veren BMNHC PH881
De beide achterpoten hebben lange veren aan onderbeen en middenvoetsbeenderen. De tenen steken buiten de veren uit. Voor de voet is dat te zien aan de linkerachterpoot (bovenste) en voor het onderbeen is het te zien aan de rechterachterpoot (bovenste).

Asymmetrie in de veren van de linkerachterpoot die onder de staart liggen BMNHC PH881
De veren van de voet zijn lang en hebben de vorm van slagpennen. Deze veren zijn duidelijk asymmetrisch, wat erop wijst dat ze van belang zijn voor vliegen.
Bij een vogel – een huidige vogel – zit het verendek van de vleugel, de voorpoot, als volgt in elkaar:

1. handpennen = primaire slagpennen = grote slagpennen
2. handdekveren
3. duimvleugel
4. armslagpennen = secundaire slagpennen = kleine slagpennen = onderarmpennen
5. grote dekveren
6. middelste dekveren
7. kleine dekveren
8. binnenste armpennen = tertiaire slagpennen = bovenarmpennen
9. schouderveren
Bij Microraptor lijkt de bevedering erg vergelijkbaar met die bij de huidige vogels. Er zijn foto’s van de veren van twee exemplaren, BMNHC PH881 en IVPP V 13352

De rechtervoorpoot van Microraptor. In de linkerfiguur is de omtrek van botten en slagpennen ingetekend. Er zijn duidelijke handpennen (primaries) en wat minder duidelijke onderarmpennnen (secundaries). BMNHC PH881
Van de rechtervoorpoot van Microraptor zijn bovenarm, spaakbeen, ellepijp en de handbotten goed zichtbaar. Er zijn drie klauwen die buiten de veren uitstaken. De hand heeft duidelijk grote slagpennen, asymmetrisch van vorm. Het aantal slagpennen is zeker groter dan zeven, en het lijkt op 10 zichtbare handpennen. Bij de huidige vogels is het aantal handpennen 9-11.
Bij een eerder gevonden exemplaar zijn de onderarmpennen duidelijker, en zijn ook dekveren zichtbaar. Hoeveel lagen dekveren er waren is niet te zien.

De rechtervoorpoot van Microraptor. Er zijn duidelijke handpennen (primaries) en duidelijke onderarmpennnen (secundaries). Ook zijn dekveren te zien (coverts). IVPP V 13352.
Kon Microraptor vliegen? Zeker: met vier vleugels. De asymmetrie in de slagpennen van voorpoot en achterpoot laat dat zien. Vliegen als bij een dubbeldekker (als dat zo heet), eerste vliegende versie van het vliegtuig.
De kleur van het verendek van Microraptor kan met goede argumenten worden weergegeven als zwart met weerschijn. Dat gaat op dezelfde manier als de kleur van Anchiornis is beargumenteerd.

Anchiornis huxleyi was grijs met zwart en witte strepen en een bruinrode kuif.
Al het zwart en grijs bij vogels is het gevolg van een zwarte kleurstof, eumelanine. Bruinig, rossig, gelig of crème is het gevolg van een roodbruine kleurstof, feomelanine.
Melanine zit in een veer opgeslagen in melanosomen. Eumelanine in eumelanosomen; feomelanine in feomelanosomen. Hoe meer melanosomen, hoe donkerder. De vorm van de melanosomen kan gezien worden met behulp van een Scanning Electron Microscope (SEM). Bij de huidige vogels hebben eumelanosomen de vorm van een afgeronde staaf; feomelanosomen zijn meer eivormig.
De melanosomen in de veren van Microraptor bleken erg lang en heel dicht op elkaar gepakt in lagen. Dat deed aan iets denken: aan weerschijn. Weerschijn is het gevolg van de structuur van een stof, met lagen waartussen lichtgolven breken en met elkaar interfereren.
Li, Vinther en anderen keken naar de melanosomen in veren van een aantal vogelsoorten. In totaal zijn er melanosomen van168 soorten bekeken, over bijna alle vogelordes.

Voorbeelden van de vorm van melanosomen. Melanosomen van (A) Noord-Amerikaanse kuifmees, Baeolophus bicolor, bruin; (B) macaroni penguin, Eudyptes chrysolophus, penguin bruin-zwart; (C) aalscholver Phalacrocorax auritus (grijs); (D) palm kakatoe, Probosciger aterrimus, zwart ; (E) Brazilie eend, Amazonetta brasiliensis, weerschijn; (F) Microraptor, monsterplaats op de schouder.

De lengte en de vorm van de melanosomen kunnen statistisch worden bewerkt en dan uitgezet in het volgende plaatje.
Statistiek op melanosoom vorm (x-as, Canonical 1) en lengte (y-as, Canonical 2). Stippen horen bij monsters van melanosomen van huidige vogelsoorten. Kleurcode: bruin = bruin, penguin = blauw, grijs = grijs, zwart = zwart, weerschijn = paars.
Nummers geven de Microraptor monsters weer. De Microraptor monsters komen terecht tussen de zwarte en weerschijn punten van de huidige vogels.
Er zijn 20 monsterpunten waar veren van Microraptor herkenbare melanosomen opleverden; de monsterpunten staan in het tweede plaatje boven aangegeven. De Microraptor monsters komen terecht tussen de zwarte en weerschijn punten van de huidige vogels.
Veren met weerschijn hebben langere en smallere melanosomen dan zwarte veren, en melanosomen voor alle andere kleuren zijn boller. Bovendien zijn melanosomen in veren met weerschijn dichter gepakt.

Tabel die de vorm van de melanosomen laat zien.
Al met al: een zwarte Microraptor met weerschijn.
Microraptor in al zijn pracht! Zwart met weerschijn.
En als de twee lange middenveren in de staart alleen bij mannetjes voorkomen, is dit een paartje:

******************
Li, Gao, Meng, Clarke, Shawkey, D’Alba, Pei, Ellison, Norell & Vinther. 2012. Reconstruction of Microraptor and the Evolution of Iridescent Plumage. Science 335:1215-1219. http://dx.doi.org/10.1126/science.1213780
Images: reconstruction by Jason Brougham; fossil photos by Mick Ellison
http://blogs.discovermagazine.com/notrocketscience/2012/03/08/a-shiny-dinosaur-%e2%80%93four-winged-microraptor-gets-colour-and-gloss/
http://en.wikipedia.org/wiki/Microraptor
http://nl.wikipedia.org/wiki/Microraptor
http://evolutiebiologie.blogspot.com/2010/02/sommige-kleuren-van-de-dino.html
http://blogs.discovermagazine.com/notrocketscience/2011/11/21/microraptor-%e2%80%93-the-four-winged-dinosaur-that-ate-birds/
http://blogs.discovermagazine.com/notrocketscience/2008/10/08/microraptor-the-dinosaur-that-flew-like-a-biplane/
http://www.jabberwocky.com/carroll/jabber/jabberwocky.html
http://nl.wikipedia.org/wiki/Iriseren